16S
rRNA 遺伝子の PCR 増幅に よ る 植物病原 MLO の検出 と 分類
東京大学農学部附属農場 なん
難 波 成 任は じ め に
マ イ コ プ ラ ズ マ 様微生物 (mycoplasma - like organ
isms ; MLO) は そ の発見以来 (土居 ら _, 1967) , 今 日 ま で約 300 以上の植物病害 の病原で あ る と 考 え ら れて き た
(McCoyet al., 1989) 。 し か し な が ら こ れ ま で, in vitro で培養で き な い こ と か ら (LEE and DAVIS, 1986) , 病気の 診断や MLO の分類 に 関 す る 研究 は な か な か進 ま ず, 専 ら 感染植物の病徴, 電子顕微鏡 に よ る 飾部組織の観察,
そ れ に 昆虫伝搬の特異性 に 関 し て研究 さ れて き た 。 し か し , こ れ ら の 実験 に は 時間がかか り , 時 に は 困難であ る た め , 多 く の MLO で そ の 性状が十分 に調べ ら れて い な い の が実状であ る 。
MLO を分類・整理す る た め に , よ り 信頼性の 高 い, 特 異的な方法 と し て , 血清試験や ハ イ ブ リ ダイ ゼ ー シ ョ ン 法の利用が考案 さ れた 。 す な わ ち , MLO に対す る 抗体 を 作出 し , ELISA や免疫電子顕微鏡法 に よ り , 感染植物 よ り MLO を 高感度 に 検 出 で き る よ う に な っ た (LIN and
CHEN,1985 ; CLARK, 1989 ; KIRKPATRICK, 1991) 。 ま た , 感 染植物 (SEARS et al., 1989 ; DAVIS et al., 1990 ; KOLLER et al., 1990 ; LEE et al., 1990 ; KUSKE et al., 1991a, b ; NAKASHIMA et al., 1992) 及 び保毒虫 (KIRKPATRICK et al.,
1987) か ら 抽出 し た MLO ゲ ノ ム の 一部 を ク ロ ー ニ ン グ し , こ れ を プ ロ ー プ と し て , MLO を植物や 昆虫 よ り 検出 し , そ の遺伝学的関係 を 調 べ た 。 こ れ ら の 方法 に よ り , 迅速で し か も 病徴 に よ る よ り も 正確 に MLO の分類がで
き る も の と 期待 さ れ る 。 し か し 一方で, MLO の生物学的 ま た は 系統分類上の位置 を知 る こ と も 重要であ る 。 最近,
16S rRNA の 研 究 が 行 わ れ る よ う に な り (LIM and SEARS, 1989 ; KIRKPATRICK et al., 1990) , 進化上, MLO は む し ろ ア コ レ プ ラ ズ マ に 近縁で, 動物マ イ コ プ ラ ズ マ と は 異 な る の で は な い か と 考 え ら れ る よ う に な っ た 。 ま た , 16S rRNA に 共通な プ ラ イ マ ー を 用 い て , MLO の 16S rRNA 遺 伝 子 を PCR 増 幅 で き る こ と や (DENG and HIRUKI, 1991) , そ の増幅 DNA の RFLP の解析 に よ り , MLO が少 な く と も 2 種類以上 に 分 け ら れ る ら し い こ と がわ か っ て き た (AHRENS and SEEMÜLER et al., 1992) 。 筆 者 ら は 最 近, 異 な る 媒 介 虫 に よ り 伝 搬 さ れ る 6 種 の MLO (表ー1) の 16S rRNA 遺伝子の塩基配列 を明 ら か に
Detection and Di任erentiation of Plant-pathogenic Mycoplasmalike Organisms (MLO) Using PCR Amplification of 16S rRNA Genes. By Shigetou
NAMBA
し , そ れ ら が 群 ( キ リ て ん ぐ 巣 (土居 ら , 1967) , ク ワ 萎縮 (土居 ら , 1967) , タ マ ネ ギ萎黄 (宮原 ら , 1982) , ト マ ト 萎黄病 MLO (加藤 ら , 1988) ) , II群 ( ツ ワ ブ キ て ん ぐ巣病 MLO (加藤 ・ 岩波, 1990) ) , 皿群 ( イ ネ 黄萎病 MLO (奈須 ら , 1967) ) の進化的 に 異 な る 三つ の ク'ル ー プ に 分 け ら れ る こ と , そ し て と く に I 群の MLO で は , 媒介 虫が異 な っ て も 進化 的 に は 同一種 と 考 え ら れ る こ と な ど が明 ら か に な っ た 。
な お , 16S rRNA の 塩基配列 に 基 づ い た 細菌分類法の 基本的 な概念 に つ い て は , 本誌前号で津 田 ら に よ り 詳説 さ れて い る の で, 本稿 で は , 16S rRNA 遺伝子 を利用 し た MLO を め ぐ る 分子遺伝学的 な 最近の研究成果 に 付 い て , 著者 ら の研究成果 を 中心 に , そ の 基礎的及 び応用 的 な側面か ら , 主 に 実験手法 を 中心 に 紹介す る こ と に し た い。 な お , 紙面が限 ら れて い る の で, 詳細 に つ い て は ま た別の機会 に 譲 る こ と と し て (S. NAMBA et al., submit
ted) , こ こ で は 専 ら 要点 の み を述べ る に と ど め た こ と を お許 し願 い た い。
1
MLO の 168 rRNA 遺伝子の PCR 増幅 に よ る 系統学的解析
1 168 rRNA の利点
地球上 に あ る 生命が l 個 の 生命体, す な わ ち 祖先生物 の子孫で あ る こ と は誰 も が認め る と こ ろ であ る が, そ の 祖先が ど ん な も の で, ど の よ う に 派生 し て き た か は よ く わ か っ て い な い 。 進化の 道筋 を 再現す る 方法 の 一 つ に , 現存す る 細胞 に あ る 分子化石 を 調べ る や り 方が あ る 。 分 子化石 と は , 系統上 き わ め て 離 れ た 生物 に 共通の構造や
表 ・ 1 本研究 で供試 し た MLO病 の 所属群 と そ の媒介虫及び地 理的発生分布
MLO群
病名 媒介虫 発生地I タ マ ネ ギ萎黄病 ヒ メ フ タ テ ン ヨコ バイ 埼玉, 兵庫, 佐賀,
ト マ ト 婆黄病 l- ー パ イ
ク ワ 萎縮病 ヒ シ モ ン ヨ コ ノ T イ ヒ シ モ ン モ ド キ キ リ て ん ぐ 巣病 |タ バ コ メ ク ラ ガ メ
ク サ ギ カ メ ム シ
H ツ 内んく 巣病
|キ マダ河 川
III イ ネ 黄萎病 ツ マ グ ロ ヨ コ バ イ
クロス ジツマグロヨコノマイ タイ ワンツマグロヨコバイ
山 口 , 長崎 佐賀, 広島 日 本, 中 国,
朝鮮半島 日 本, 中 国 , 朝鮮半島, 台湾 宮崎
日 本, 東南 ア ジ ア
一一一
38
一一一16S rRNA 遺伝子 の PCR増幅 に よ る 植物病原MLOの検出 と 分類
87 機能 を 指 し , お そ ら く 最初の細胞 に 既 に 存在 し て い た と
考 え ら れ る も の であ る 。 中 で も 遺伝子 の変異の集積 は極 め て遅 く , 保存性の 高 い RNA や タ ンパ ク 質 を コ ー ド し て い る 遺伝子 に 集積 し た 変化 (変異) の 数 を 数 え る と , 生物相互の進化上の 類縁関係 を 推測 す る こ と がで き る と す る 考 え 方が あ る 。 共通の祖先か ら 始 ま っ た 生物の進化 の過程 は 系統樹 と し て 表す こ と がで き る 。 こ れ ま でグ ロ ビ ン や シ ト ロ ク ロ ム c な ど の よ く 保存 さ れて い る タ ンパ ク 質や遺伝子が分子時計 と し て利用 さ れて き た 。 し か し 微生物の 中 に は シ ト ク ロ ム c を 欠 く も の が あ り , ま た仮 に 持 っ て い て も 相互の タ ンパ ク 質が違い す ぎ て , 類縁関 係 を確定 し に く い も の が あ る こ と か ら , MOESE ら (1980) は 16S rRNA の利用 を 試 み た 。 そ の利点 は , ① 生物 に 普 遍 的 に 存在 し , そ の構造 と 機能が よ く 保存 さ れ, 系統樹 の作成が比較的容易 で あ り , ② 大量 に 存在 し , 精製 ・ 分 析が し や す い事 に あ る 。 今 日 , 16S rRNA の塩基配列 を 分子化石 と し て 用 い る こ と に よ り 得 ら れ た 結果 は 強 く 支 持 さ れて お り , そ の 意味で, 16S rRNA 遺伝子 を 用 い た 系統発生学的 な比較研究 は , Mollicutes 網の微生物 と 考 え ら れて い る 培養 の 困 難 な MLO の進化学的 な 由来 と 生 物分類上の位置づ け に , き わ め て 有効 な 武器 と な る 。
2 Molllicutes 綱の分類学的位置
Mollicutes 綱の細菌 は ヒ ト や動物, 昆虫, 植物の 重要 病害 に 関連 し た も の が多 く (RAZIN and BARILE, 1985) , 細 胞壁 を 欠 き , 小型であ る 点 で, 他の 原核微生物 と は 区別 さ れ る (表 2) 。 事実, そ れ ら は最小の 自 己複製微生物で あ る と 考 え ら れて い る 。 動物マ イ コ プ ラ ズ、マ の ゲ ノ ム は,
約 680-1600 kb で (NEIMARK and LANGE, 1990) , こ れ は , 大腸菌 の 4700 kb や 植 物 の 6000-60000 Mb に 比 べ る と 格段 に 小 さ し ミ 。 こ れ ま での研究 で は , 16S rRNA 及び 5S rRNA の 塩基配列, ま た 16S rRNA オ リ ゴ ヌ ク レ オ チ ド カ タ ロ グ の解析な ど に よ り , Mollicutes 綱の細菌 は 系 統学 的 に は グ ラ ム 陽性細菌 に 近縁 で, cIostridia か ら 退 化的な進化 を 遂 げて 発生 し て き た も の と 考 え ら れて い る (WOESE et al., 1980 ; MANILOFF, 1983 ; R∞ERS et al.,
1985 ; WEISBURG et al., 1989) 0 rRNA の配列が似て い る こ と に加 え て , Mollicutes 綱 は ゲ ノ ム の G + C 含量の低
表 -2 Mollicutes 綱 の 分類
K恥... ・ Dlvlalon 回a岨 0"'.. Famlly GenU8
い点で も cIostridia に 似 て い る 。
3 MLO の 168 rRNA 遺伝子の PCR に よ る 増幅
16S rRNA の解析 は 従来, 逆転写酵素 を 利用 し た り , ゲ ノ ム ラ イ ブ ラ リ ー を 作製 し て 行わ れて い た が, 既 に 述 べた よ う に , こ の遺伝子 は微生物 を 通 じ て 高度 に 保存 さ れて お り , ま た Mollicutes の 多 く でゲ ノ ム に 約 1-2 コ ピ ー存在す る 事が知 ら れて い る (SAWADA et al., 1981) 。 そ こ で ま ず, Mollicutes の 16S rRNA 遺伝子 を デ ー タ ベ ー ス を 用 い て 解析 し , MolIicutes 網及 び リ ケ ッ チ ア 類 に 特異的 な プ ラ イ マ ー を デザ イ ン ・ 合成 し た ( ユ ニバ ー サ lレ プ ラ イ マ ー セ ッ ト , 図-1) (SOMMER and TAUTZ, 1989) 。 次 に MLO 濃縮分画試料 を LEE and DAVIS (1988) の 方法 に基づ き 若干の改良 を 加 え て , 酵素処理 に よ り 調製 し た 。 PCR 反応 は以下の条件で行 っ た 。 各抽出 DNA サ ン プル 1 μl (MLO-濃縮分画 DNA (収量 6 μg/g 維管束組織) : 約 8 ng ; Mycoplasma orale 抽 出 全 DNA) を PCR 反応 の鋳型 と し て 用 い た 。 増幅反応 は , 20 μl PCR 反応 ミ ッ ク ス (各 250 μ M の dATP, dCTP, dGTP, dTTP ; 各 1 μ M5'- 及 び 3' プ ラ イ マ ー ; PCR 反 応ノ T ツ フ ァ ー : 0 . 5 U Taq DNA ポ リ メ ラ ー ゼ ( プ ロ メ ガ, USA な ど) ,
100 μJ 流動パ ラ フ ィ ン (No. 261-32GR, ナ カ ラ イ テ ス ク で十分で あ る ) ) に よ り 行 っ た 。 シ ー ク エ ン ス 用 の 鋳型調 製 は 100 μJ の ス ケ ー ル の PCR 反応で行 っ た (NAMBA et al., submitted) 0 PCR 反応 は サ ー モ プ ロ セ ッ サ TR-100
( タ イ テ ク (株) ) を 用 い て , 特 に 断 ら な い 限 り 50 サ イ ク ル行 っ た 。 サ イ ク ル条件 は , 以下の 通 り で あ る : 最初の サ イ ク ル : 熱変性 (94 0C, 90 秒間) , 温度降下 (40 秒間) , ア ニ ー リ ン グ (60 0C, 2 分間) , 温度上昇 (30 秒間) , 伸 長反応 (72 oC, 3 分間) ; 続 く 50 サ イ ク Jレ は 熱変性 を 2 分間で行 う 他 は ア ニ ー リ ン グ ・ 伸長反応 は 最初の サ イ ク ル と 同 じ ; 最後 の 伸長 反応 は 7 分 間 行 う (SAIKI et al.,
1988 ; INNls and GELFAND, 1990) 。 以下, 本稿で は 特 に 断 ら な い 限 り こ れ を 「通常 の 条件J と 称 し た 。
本研究 で は 、 各種の媒介虫 に よ り 伝搬 さ れ る 6 種類の
プライマー
Unlversal p同mer8et
サイズ (bp)
1 6S rRNA 遺伝子
1370 ・・-
MlO specltlc 750 睡E・E・-
咽時ー園田園ー
MLO-Ia帥eiflc 世間 4‘E・E・-
ULO-U apeclßc 叩70 唾E・E・-
UlO-lIlSpeclfic 350 ...-
図 - 1 各 プ ラ イ マ ー の位置 と 鳩幅 DNA の サ イ ズ
一一一
39 一一一
MLO 病 に つ い て そ れ ぞれ PCR 増 幅 を 試 み た 結果, そ の い ずれか ら も 予想 ど お り 約 1400 bp の DNA 断片が増幅 さ れた 。 こ の DNA は , 健全試料か ら は 増幅 さ れず (図ー 2) , ま た サ ザ ン及 び ノ ー ザ ン ハ イ プ リ ダイ ゼ ー シ ョ ン に よ り , MLO 感染植物 に 特異的で, 16S rRNA 遺伝子 に相 補的な配列 を持つ こ と が確認 さ れた 。
4
PCR 増幅 DNA の塩基配列決定
各種生物の 16S 及 び 16S 様 rRNA の塩基配列 の デ ー タ を デー タ ベ ー ス よ り 検索 ・ 解析 し (NEEFS et al., 1990) , 18 種の シ ー ク エ ン ス 用 プ ラ イ マ ー を 合成 し た 。
各 MLO サ ン プル よ り PCR 増幅 し た 反 応液 を精製 し て シ ー ク エ ン ス 反応の鋳型 と し , 合 成 プ ラ イ マ ー を 用 い て ダイ レ ク ト シ ー ク エ ン シ ン グ に よ り 塩基配列 を 決定 し た。 シ ー ク エ ン ス 反 応 は, 35S dATP を 用 い て Se-
1 2 3 4 5 6 7 8 9 1 0 1 1 12 13 14 15
“ 4・E
図 - 2 ( l )
MLO 濃縮分画試料 ま た は 全 ONA 試料か ら ユ ニ パ ー サ ノレ プ ラ イ マ ー を 用 い て 増幅 し た ゲ ノレ泳動写 真 (矢印上 1370 bp; 下 :
750 bp) (H : 健全試料 ; 0 : そ れぞれの MLO に 感染 し た 試料)l λ Hind III
•
EcoRI マ ー カ ー2 タ マ ネ ギ (H ) (MLO 濃縮分画法 に よ り 調製 し た 試 料 を PCR)
3 タ マ ネ ギ (0) (MLO 濃縮分画 よ り : 以下の レ ー ン は全 ONA 試料)
4 タ マ ネ ギ ( H ) 5 タ マ ネ ギ (0) 6 ツ ワ プ キ ( H ) 7 ツ ワ プ キ (0)
8 イ ネ (H )
9 イ ネ (0)
10 Mycoþlasma orale
1 4 無 毒 媒 介 虫 ( ヒ メ フ タ テ ン ヨ コ バ イ ) の 全 ONA 試料
15 保毒虫
(2) 全 ON A 試料か ら ユ ニ バ ー サ ル プ ラ イ マ ー セ ツ ト 及 び MLO に 特 異 的 な プ ラ イ 7 ー を 用 い て RPCR 法 に よ り 増 幅 し た ONA の ゲ ノレ泳動写 真
11 タ マ ネ ギ ( H ) 12 タ 7 ネ ギ (0) 13 MycoþよI1Sma orale
quenase Version 2.0 (USB, USA) に よ り , 図 -3 の プ ロ ト コ ー ル に 従 っ て 行 っ た (TABOR and
RICHARDSON,1989 ; NAMBA et al., submitted) 。 そ の 結果, 1370 塩基 の 配 列 が 決 定 さ れ, そ れ ら は い ず れ も Mollicutes の 16S rRNA と 高 い相向性 を 示 し た 。 ま た そ れ ら は , そ の塩基 配列の相向性か ら 表 l の よ う に 三 つ の 群 に 分 か れ た 。 I 群の 本邦産 MLO の配列 は 全 て 同 じ で あ っ た 。 ま た 米国 で報告 さ れた western aster yellows MLO の 強毒系統 (SAY-MLO) と Oenothera hookeri の フ ィ ロ デ ィ ー 症状 に 関与す る MLO (O-MLO) o ( 1 群 と の相 向性 は そ れぞ れ 99 . 7%及 び 99 . 8% ) は I 群 と 相向性が高か っ た 。
本研究 で用 い た 合成 プ ラ イ マ ー は, ス ピ ロ プ ラ ズ マ
(WIIITCO�IIl,1 980) に も 共通 な 配列 を も つ が, シ ー ク エ ン ス の 結果, 増幅 さ れ た DNA に は ス ピ ロ プ ラ ズ マ に ホ モ ロ ジ ー の あ る 配列 は認 め ら れ な か っ た 。
MLO の 16S rRNA の G + C 含量 は , 動物 マ イ コ プ ラ
ズ マ を は じ め と す る Mollicut白 の そ れ と ほ ぽ同 じ で, 大 腸菌 な ど他の細菌 に比べ, 非常 に 低 い事が知 ら れて い る が, 本邦産 の 6 種の M LO で も , 同 様 に 低 い レ ベ ル で あ っ
た。こ れ ら I �III 群 の 配 列 デ ー タ を 用 い て , ま ず Gen
Bank デ ー タ ベ ー ス に 登録 さ れ て い る す べ て の 生 物 の rRNA ( 16S ま た は 18S) の 配列 と 相 向性の比較 を 行 っ た 。 そ の 結果, 比較的近縁で あ る こ と が明 ら か に な っ た 属の細菌 を 中 心 に 8 属 の 細菌, す な わ ち :
E.coli ( グ ラ ム 陰性細菌) , Bacillus subtilis ( グ ラ ム 陽性細菌) , Metha
nococcLls vannielii (古 細 菌) 及 び Mollicutes 綱 の A今copl,αsma gallisψticum (動物マ イ コ プ ラ ズ マ ) , Urea-
シークェンスプライマーの アニー リ ング
在日
!�7';j"",Ò/日
E
アニー リ ン : 水 4 μ I
グチューブ 反応以 ッ フ ァ 2 プラ イ マー 2 D N A副業
9 9 5 0 C ( 5 分) ー氷冷 ( 1 0分)
D T T μi
ラ ベ リ ングミ ク ス 2 35 S d A T P 0 . 7 マ ンガンバッ フ ァ 0 . 7 加盟r田晴釈液 2
6 . 4 3 . 5 μ I ずつ分注
ア ナ ロ グチュープ (予め各2 . 5 μ J の ア ナ ロ グ ミ ッ クスを分注し 3 7 0C保温)
|一
一 (各4 . 5 μ 1 )9 0 0 C ( 5 分) →氷冷 シ ー クェンスゲルに 3 μ l ずつ アプライ したのち泳動する
図 - 3 シ ー ク エ ン ス 反 応の手順
一一一
40一一一
89
明 ら か に す る 目 的で, 配列 デ ー タ を 元 に , 系統樹 を 作成
し た 。 進化距灘 (KIMURA. 1980) を Knuc 値 (塩基置換率) に よ り 評価 し , Neighbor joining (NJ) 法 (近隣結合法) に よ り (SAlTOU and NEI. 1987) 系統関係 を 解析 し て 系統 樹 を 作成 し た ( 図-5) 。 基準菌 に は , Methanococcus van
nielii を 用 い た 。 系統樹の 各横輸 の 長 さ は Knuc 値 の 大 き さ (系統進化上の 隔 た り ) を 示 し て い る ( ス ケ ー ル参 照) 。 ま た こ の 系 統 樹 は , 分 子 進 化 学 解 析 シ ス テ ム rODENJ を 用 い た ブ ー ト ス ト ラ ッ プ 法 (FELSENSTEIN.
1985) に よ り 評価 し た ク ラ ス タ ー ト ポ ロ ジ ー の 信頼度 (% ) か ら , 比較的信頼性の 高 い も の で あ る 事が確認 さ れ た 。 こ れ を 見 る と , 米 国 の MLO (O - MLO と SAY MLO) は そ れ ぞ れ I 群の MLO に 含 ま れ る こ と を 示 し て お り , こ の こ と は , 地理的 に 隔離 さ れた あ る 種の MLO が 同様な祖先か ら 進化 し て き た 事 を 示唆す る も の で興味深 いo Knuc 値か ら , MLO や動物マ イ コ プ ラ ズ マ で は , R subtilis よ り も 進化の速度が大 き し ま た MLO の I I 群 と III群 は比較的近縁で あ り , 両群 と も I 群 よ り も 進化速度 が大 き い も の と 推定 さ れ る 。 MLO は ア コ レ プ ラ ズ マ と ア ナ エ ロ プ ラ ズ マ に 近縁で あ る が, ス ピ ロ プ ラ ズ マ , 動 物マ イ コ プ ラ ズ マ , ウ レ ア プ ラ ズ マ と は別 の ク ラ ス タ ー を形成 し て い る 。 こ の こ と は , MLO が Mollicutes 綱の メ ンバ ー の マ イ コ プ ラ ズ マ や ス ピ ロ プ ラ ズ マ , ウ レ ア プ ラ ズ マ か ら , 早期 に 分 か れ て き た こ と を 示唆 し て い る 。
こ の よ う に , MLO は 動物 マ イ コ プ ラ ズ マ や (S citri や S kunkelii の よ う な ) 植物病原 ス ピ ロ プ ラ ズ マ と は 進化 的 に 異 な る と と も に , そ れ 自 身変異 を も っ た ク' ル ー プで あ る こ と が明 ら か に な っ た 。 ま た , 1 群の 本邦産 MLO の 塩基配列 に 変異が認め ら れ な い こ と は , 進化的 に 保存 さ
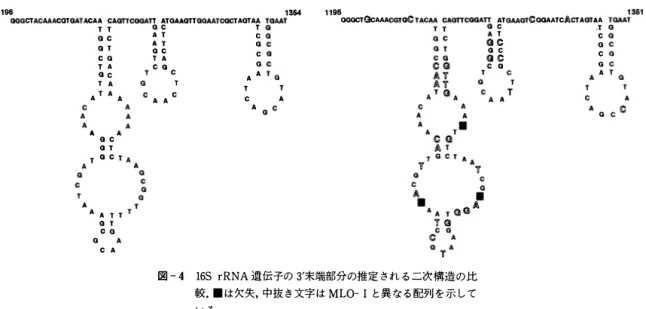
16S rRNA遺伝子 の PCR増幅 に よ る 植物病原MLOの検出 と 分類ρL俗ma urealyticum ( ウ レ ア プ ラ ズ マ ) , Acholeþlasma laidlaωii ( ア コ レ プ ラ ズ マ ) , Spiropゐsma citri ( ス ピ ロ プ ラ ズ マ ) , Anaeroplasma varium ( ア ナ エ ロ プ ラ ズ マ ) , 及び米国の MLO 2 種 : SAY-MLO, 0一MLO の そ れ ぞ れ と 詳細 に比較 を行 っ た . MLO と Mollicutes 綱の配列 を比較す る と , E coli, B. subtilis に は な い デ リ ー シ ョ ン がい く つ か共通 し て 認め ら れ る 。 ま た , こ れ ら の細菌 を 通 じ て 高度 に 保存 さ れた 領域 (保存領域) と 異 な る 領域 (可変領域) が認め ら れ る 。 し か し な が ら , こ れ ら の 可変 領域 も , そ の二次構造 を比較す る と 極 め て よ く 似 て い る
(GUTTEL et al.. 1985) ( 図-4)
各 MLO と Mollicutes 綱 の 各属 細 菌 に つ い て ユ ニ ー ク な オ リ ゴ ヌ ク レ オ チ ド の 有無 に つ い て 調べ て み る と , MLO は ア コ レ プ ラ ズ マ や ア ナ エ ロ プ ラ ズ マ に 近縁であ
る が, 動物マ イ コ プ ラ ズ マ や ス ピ ロ プ ラ ズ マ , ウ レ ア プ ラ ズ マ と は進化 的 に 離れて お り , 両者が早 い ス ピ ー ド で 進 化 的 変 遷 を 遂 げ た と 推 測 さ れ る (表 ー3 ; LIM and SEARS. 1989 ; WOESE et al.. 1980) 。 ま た SAY-MLO と O
MLO の オ リ ゴ ヌ ク レ オ チ ド パ タ ー ン は い ずれ も MLO I 群 と 全 く 一致 し て い た 。 WOESE ら (1980) は , Mollicutes 網の細菌で は , す べ て の真正細菌 の 16S rRNA で認 め ら れ る 高度 に保存 さ れた配列 の 多 く が欠失 し , そ の代わ り に 多 く の ユ ニ ー ク な配列が認 め ら れ る こ と か ら , そ の 高 い 突然変異率が結果 的 に 急速 な 進化 を も た ら し た も の と 説明 し て い る 。 実際 こ の ユ ニ ー ク な配列 は , 各 MLO 群 に 認め ら れ る が, Mollicutes 綱の 多 く の 属 に認め ら れ る 保 存配列 も 欠失 し て い る 。
5 MLO の系統学的 な 比較
Mollicutes 綱 の 他 の 属 と MLO の 系 統 学 的 な 関 係 を
Mycoρlasma 伊111..ρ血um
M 民地 甜•• ,J
e--dC HGGCGCT aτCGCGA c G
A Y AA m咽
c 町剖 A 町A C Y 叫 T-pu MCYCCCG CTT naAG@Gcr A 3・・ =A
刷 1
GE Tee-A
白幽 ・組問 W
AA. A A円 剛山 A"?冒 T, ,町、俺崎 帥YCT@GTT G
GTC YGOAAr 凶YGGC岳AAY CAG ATeEa
白 A
AY A {
W
AA C
TBa. cc,a
ω ・・凶刷ι国 一s のMg
MLO・1
nGGC@CV 凶 Gc '・a-同 M s ・-ra偶
MTC@CG O AA 町 AYC AMR V 叩T a a-【個 叩a-R 倒
町CYYCAG CTC A T-aUA民・"のu'・ph aMR 刷 YGC Gcee
G Ay m
AAA A
T
g A
Aτ Y
A
MYCTGACA A CYC TYGA AYGGCTey y a@G
Aecc 帥 AA τ AG
冶
C ‘A
AA
馴 闇輔' 叩A." 周.掴偶R A" F』切' a" 町n ecy d
図 - 4 16S rRNA 遺伝子の 3'末端部分の推定 さ れ る こ次構造 の 比 較. . は 欠失, 中 抜 き 文字 は MLO- 1 と 異 な る 配列 を 示 し て
い る .
一- 41 一一一
( 1993 年) 第2号
疫 第47巻 防
植 物 90
Mollicutes 網の 各種の 16S rRNA 遺伝子 に お け る ユ ニ ー ク な オ リ ゴ ヌ ク レ オ チ ド の存在
occurをlIlC掴 10
•
表 - 3
Se哩lSlC・
MCD
= r::ÞN>l>>岨
�岨 MUlJ(J(D N.1ÞCXllI園 田JI\I'a且且 UI温xx丈盟抵酒 田圃JPCIØ喧 MIlUXXX>\田直B
E栓S申ÙIæ1/l othsr
�yticun bactarla
{ ・ }ω
+
+ +
・
・ + +
・ +
+
h官官司:>1"",国 AC:b>I司:>1aæa 皐ir可証aBI1歯 周悼記事立...
四r:i四 laidJ.awii cl包r1 c唱;Ir1col国
+ + + +
・ + +
・ +
+
+ + + +
・ + +
・ +
+
+ + 同
・ +
・ + + +
・
+
・
・ + +
・
・
・
・
・
殴I[D・I:tI
+ +
・ + +
・
・
・ +
・
MIJ:)-:tI
+ +
・ + +
・ 司
・
・
・
MU)-工
+ +
・ ・ +
・ +
・ +
・
Q-MI!) S町ー匹。
+ +
・
・ +
・ +
・ +
・
ろ , phytoplasma ( フ ァ イ ト プ ラ ズ マ ; phyto- ( ギ リ シ ャ 語 (phuton) で 「植物」 の 意) + -plasma ( ギ リ シ ャ 語 (plasma) で 「形成 さ れた も の 」 の意) ) の よ う な 呼称 の 方 が適 当 で あ る と 考 え ら れ る よ う に な っ て き た 。 ま た こ れ ら MLO の 系統発生学的な比較か ら , 少 な く と も I ・ II ' III の複数の 群 に 類別 さ れ る 進化的 に 異 な っ た グル ー プか
ら な る こ と , ま た , 媒介昆虫が異な り , 地理的 に 隔離 さ れた地域 に 発生す る MLO で も , 系統発生学的 に は ほ ぽ 同ーの MLO か ら な る 群 ( I 群 MLO) が存在す る こ と が 明 ら か に な っ た 。 以上 の 知見 は , 培養の 困難 な MLO の人 工培養 に 手がか り と な る も の で あ り , ま た MLO の 検 出 診断法 に 新た な 可能性 を 示唆す る も の で あ る (NAMBA et al., submitted) 。
MLO-Il
Mycoplasmø gatllseptJcum 島創出lus subfJlIs
a)非常にま れ.
u,閣内酬叫刷Iyt蜘m Splroplasma cnrl
図 -5 Mollicutes 綱 の 各属細菌の 16S rRNA 遺伝子 の 配 列 に 基づ い て 作成 し た 系統樹. 横棒の長 さ は 系統進 化 的 な 隔 た り を 示す.
0.01 Kn国
168 rRNA 遺伝子の PCR 増幅 に よ る MLO の検出
E
Mollicutes 綱 の 細 菌 の 高 度 に 保存 さ れ た rRNA (5S 及び 16S) の塩基配列 や オ リ ゴ ヌ ク レ オ チ ド 分類 に 関 す る 研究 に よ り , Mollicutes の起源や 系統発生学的 な 関係 が解明 さ れて き た (WOESE et al., 1980 ; MANIむOFF, 1983 ; WOESE, 1987 ; WEISBURG et al., 1989) 。 ク ロ ー ン化 し た rRNA 遺伝子 を プ ロ ー プ に 用 い て , 細胞培養 に お け る マ イ コ プ ラ ズ マ の コ ン タ ミ ネ ー シ ョ ン を検出 し た り (GOBEL and STANBRIDGE, 1987 ; HAIlASAWA et al., 1986) , AIDS 患 者か ら の マ イ コ プ ラ ズ マ の検出 江ρ et al., 1989) , さ ら に Mollicutes の 種 レ ベ ル で の 検 出 な ど が 試 み ら れ て き た (GOBEL et al., 1984) 。 ま た , マ イ コ プ ラ ズ マ の 16S rRNA 遺伝子 に 高 い ホ モ ロ ジ ー を 持 つ オ リ ゴ ヌ ク レ オ チ ド プ ロ ー プ に よ り , MLO の検出 は ク ロ ー ン 化 し た ク ロ モ ゾー ム DNA プ ロ ー プ よ り も 1O�20 倍 も 検出 感度 が向上 し た (KIRKPATRICK and FRASER, 1988) 。 最近, マ イ コ プ ラ ズ マ の コ ン タ ミ ネ ー シ ョ ン が PCR に よ り 検出 で
一一一
42 一一一 れ た 均 一 な 群 を 形 成 し て お り , 異 な る 媒 介 虫 を も っ
MLO よ り な る 群 に も 関わ ら ず, MLO と し て は進化的 に き わ め て近接 し た ス テ ー ジ に あ る も の と 考 え ら れ る 。 ま た , II ・ III群の MLO も そ れぞれ固有の異 な る 媒介虫 に よ り 伝搬 さ れ, 特 に , II群の媒介虫 は I 群の MLO を媒介 し な い し , そ の逆 も 同様であ る (加藤 ・ 岩波, 1990 ; 加藤 ら et al., 1988) 。 以上の こ と は, 媒介虫特異性が, 比較的新 し い進化的適応現象であ る 事 を 示唆 し て い る 。 し か し な が ら , MLO は媒介虫体内 で増殖す る わ け で, MLO と 媒 介虫 の親和性 は や は り MLO 分類上重要 な フ ァ ク タ ー で
あ る と い わ ね ば な ら な い だ ろ う 。
MLO は 培養 に 成功 し て お ら ず, ま だ分類上の結論は 得 ら れて い な し 当 。 し か し な が ら , 以上の デー タ よ り MLO は Mollicutes 綱 に 分類す る の が適当 で あ り , ま た , マ イ コ プ ラ ズ マ ・ ス ピ ロ プ ラ ズ マ ・ ウ レ ア プ ラ ズ マ に比べ,
ア コ レ プ ラ ズ マ や ア ナ エ ロ プ ラ ズ マ に よ り 近縁であ る と
考 え ら れ る 。 ま た こ の こ と は, 従来 こ の微生物が MLO と
し て 呼称 さ れて き た こ と に 再考 を促す も の で あ り , む し
16S rRNA遺伝子の PCR増幅 に よ る 植物病原MLOの検出 と 分類 91
き る こ と も 報告 さ れて い る (KUNlTA et al., 1989) 。 さ ら
に PCR に よ り , MLO ゲ ノ ム DNA の精製試料か ら , 16S rRNA 遺伝子 を 特異 的 に 増幅 で き る こ と も 明 ら か に な
っ た (DENG and HIRUKI, 1991) 。
本項で は , 権病植物 よ り 直接抽出 し た 全 DNA か ら も ・ 効率 的 に MLO の 16S rRNA 遺伝子 の み が増幅で き る
こ と , ま た , MLO の 各群 を 判別 す る 特異的な プ ラ イ マ ー を合成す る こ と に よ り , そ れ ぞれの グ ル ー プ を 特異的 に 増幅で き る こ と , 及 び微量の DNA サ ン プル よ り , MLO か ど う か, さ ら に そ の所属す る 群 を判別す る 一連の PCR 反応 を , 1 本 の チ ュ ー プ で し か も 短 時 間 行 う 「二段 階 PCR 法」 を 紹介す る (NAMBA et al., Submitted) 。
1 PCR 増幅 に用 い る 鋳型 DNA の調製
植物体か ら 抽 出 す る PCR 増幅用 の 鋳型 DNA は, 既 報の 方法 (KOLLAR et al., 1990 ; NAKASHIMA et al., 1991) を 一部 改 変 し て , 植 物 体 の 各 部 位 の d躍 病 組 織 よ り 全 DNA を調製 し て得 た 。 す な わ ち , 各 0 . 3 g の植物組織 を 液体窒索中 で磨砕 し , 融解後, 0 . 9 ml の抽出パ ッ フ ァ ー (1 . 4 M NaCl, 1 % PVP, 2 % CTAB, 0 . 1 M Tris-HCl (pH 8 . 0) , 20 mM EDTA (pH 8 . 0) ) を 加 え , 乳鉢で磨 砕 い 1 . 5 ml の遠心チ ュ ー ブで 65 .C, 30 分 イ ン キ ュ ベ ー ト し た後, 1 , 200 g で 5 分遠心す る 。 遠心後のよ滑 に 等 量の ク ロ ロ ホ ル ム ー イ ソ ア ミ ル ア ル コ ー ル (24 : 1) を加 え抽出 し , 1 , 200 g で 5 分遠心後, 上清に 0 . 7 vol 以下の イ ソ プ ロ パ ノ ー ル を 加 え ボ ル テ ッ ク ス 後, 室温で 5 分静 置 し , 同様 に 遠心す る 。 沈澱 を 50 μl の TE (10 mM Tris -HCl (pH 8 . 0) , 1 mM EDTA (pH 8 . 0) , pH 8 . 0) で懸 濁 し , PCR 反応の鋳型用 DNA サ ン プル と す る 。
2 プ ラ イ マ ー お よ び PCR 増幅 に よ る MLO の検出
塩基配列 の デ ー タ を も と に し て , MLO の 各群 を判別 で き る よ う な特異的 な プ ラ イ マ ー を デ ザ イ ン し た (図 1) 。 す な わ ち , (A) Mollicutes 網 に 特異的でか つ そ の 中 の各微生物 に共通 な 5'ー及 び 3' プ ラ イ マ ー ( ユ ニ パ ー サ ル プ ラ イ マ ー ) セ ッ ト パB) MLO に特異的な プ ラ イ マ ー (以下 3'ー プ ラ イ マ ー ) , (C) 1 群 に 特異 的 な プ ラ イ マ ー,
(D) III群 に 特異 的 な プ ラ イ マ ー を そ れ ぞ れ合成 し た 。 PCR 反応の条件 は , ア ニ ー リ ン グ温度 を 40 .Cで行 っ た 他 は , プ ラ イ マ ー (C) 及 び (D) に は通常の条件で 10 サ イ ク ル, プ ラ イ マ ー (B) に は 同様 に 20 サ イ ク ル行 っ た 。 い ずれ も 5'ー プ ラ イ マ ー に は プ ラ イ マ ー セ ッ ト (A) の 5'ー プ ラ イ マ ー を 用 い た 。
ユ ニ バ ー サ ル プ ラ イ マ ー セ ッ ト を 用 い た PCR 反 応 に よ り , 各種感染植物, 保毒媒介昆虫及び動物マ イ コ プ ラ ズ マ 培養菌体か ら 抽 出 し た 全 DNA 試料か ら , ほ ぽ全長 の 16S rRNA 遺伝子の DNA が増幅 さ れ た ( 図 ー2) 。 ま た , MLO, 1 群及びIII群の お の お の に 特異的 な プ ラ イ マ ー セ ッ ト を 用 い た と こ ろ , 予想 さ れた サ イ ズ の DNA 断
片 が そ れ ぞ れ増幅 さ れた ( 図 -6) 。
3 二段階 PCR ( Recycled-PCR (RPCR) ) 法
各 プ ラ イ マ ー を 用 い て PCR に よ る 検 出 法 の 改変 を 試 み た (NAMBA et al., Submitted) 。 す な わ ち , ま ずユ ニバ ー サ ル プ ラ イ マ ー セ ッ ト を 用 い て PCR 反 応 を 行 い , 増 幅 さ れ た DNA に つ い て , さ ら に そ れ が MLO の 16S rRNA で あ る か ど う か を PCR 反 応 に よ り 確認 し た い わ け で あ る が, そ の つ ど PCR 反 応 を 行 っ て い る と , 時間 と 手間及 び経費が か か る 。 そ こ で最初 に PCR 反 応 を 行 っ た チ ュ ー プ に 適 当 な プ ラ イ マ ー を加 え る だ け で, 他 に は 何 も 加 え ず に PCR 反応 を 継続 し て行 っ た 。 す な わ ち , ユ ニバ ー サ ル プ ラ イ マ ー セ ッ ト に よ り 16S rRNA 遺伝子 部分 を 十分量 PCR 増幅 し (第 1 段階) , そ こ に 内 部 プ ラ イ マ ー (B, C ま た は D) を 添加 し , PCR 反応 (第 2 段 階) を行 っ た ( 図ー7) 。 サ イ ク ル条件 は , 第 1 段階 は 通常 の条件で 10 サ イ ク ル行 っ た 。 第 2 段階 は , ア ニ ー リ ン グ 温度 を 40 .C に し た 他 は , 通常 の 条件 で 20 サ イ ク ル行 っ た 。 我々 は こ の PCR 増幅法 を 「二段階 PCR (Recyc1ed -PCR (RPCR) ) 法」 と 称 し た 。
RPCR に よ り , MLO 特異的 な 16S rRNA 遺伝子 の 断 片が非常 に 効率 よ く 増幅 さ れ る 。 最 近 DENG and HIRUKI
(1991) は , ア ニ ー リ ン グ 温度が低 い (50 .C以下) と , 健 全植物や E. coli や ク ロ ロ プ ラ ス ト の 16S rRNA 遺伝子 は Mollicutes 綱の 16S rRNA と 高 い ホ モ ロ ジ ー を 持 っ て い る の で, 非特異的 な DNA 増幅が起 こ る こ と を 報告 し て い る 。 本研究 で新た に 開発 し た RPCR 法 は , こ の 非 特異反応 を効果 的 に 抑 え る こ と が出来た ( 図-6) 。 こ の 二 度 の PCR 反応 に よ り 得 ら れ る 増 幅 DNA は , ア ガ ロ ー ス 電気泳動 に よ り , 第 l 段階及び第 2 段階で そ れ ぞ れ増 幅 さ れ る DNA バ ン ド の複合ノ T タ ー ン と し て 得 ら れ る 。 RPCR 法 に よ り , MLO に 特異 的 な プ ラ イ マ ー を 用 い て , 各群の 16S rRNA フ ラ グ メ ン ト が, 植物体及 び媒介虫体 内か ら 効果 的 に 増幅 さ れ (NAMBA et al., submitted) , こ れ に よ り , MLO の 所属群 を 効率的 に 判別 で き る こ と が わ か っ た (図-2, 6) 。
4
媒介虫 か ら の MLO DNA の検出タ マ ネ ギ萎黄病 MLO の媒介虫 で あ る ヒ メ フ タ テ ン ヨ コ パ イ を 5 頭用 い て そ の 磨砕組織 よ り 全 DNA を 抽 出 し ユ ニ バ ー サ ル プ ラ イ マ ー セ ッ ト を 用 い て PCR を 行 っ た と こ ろ , 無毒虫及 び保毒虫 の双方か ら , タ マ ネ ギ萎黄病 MLO 擢病株 よ り 増幅 さ れ た の と 同 サ イ ズ の DNA が増 幅 さ れた ( 図-2) 。 そ こ で MLO に 特異 的 な プ ラ イ マ ー を 内部 プ ラ イ マ ー に 用 い て RPCR を 行 っ た と こ ろ , 1370 bp のバ ン ド は無毒 ・ 保毒虫 の両者 よ り 検出 さ れた が, 730 bp のバ ン ド は保毒虫か ら の み増幅 さ れた ( 図 -6) 0 1370 bp の バ ン ド は ユ ニ バ ー サ ル プ ラ イ マ ー セ ッ ト に よ り 増 幅 さ れ た バ ン ド で あ る 。 保 毒 虫 の 730 bp バ ン ド は ,
一- 43 一一一
RPCR の 二段階 自 に 加 え た MLO 特異 的 な プ ラ イ マ ー (B) と 最初 に 加 え た ユ ニ バー サ ル プ ラ イ マ ー セ ッ ト の 5' ー プ ラ イ マ ー に よ り 増幅 さ れ た DNA と 考 え ら れ る 。 実 際, 1370 bp と 730 bp の バ ン ド を ゲ ル よ り 再抽 出 し , そ
れ ぞ れ シ ー ク エ ン ス し た と こ ろ , I 群の 1 6S rRNA と それぞれ配列が一致 し た 。 し か し , 無毒虫 の 1370 bp 及 び 900 bp の バ ン ド は , MLO の I �III群 の い ずれの配列 と
も 全 く 異 な り , リ ケ ッ チ ア や ク ラ ミ ジ ア に 最 も 相向性が 高か っ た 。 し た が っ て こ の DNA は, MLO 病 に 関与す る バ ク テ リ ア 由来の も の で は な い と 考 え ら れ る 。 そ こ で,
MLO に特異的 な プ ラ イ マ ー セ ッ ト ( (B) 及 びユ ニ パ ー サ ル プ ラ イ マ ー セ ッ ト の 5' プ ラ イ マ ー ) を 用 い て 直接両サ ン プlレ よ り PCR を 行 っ た と こ ろ ( 図 6) , 保毒虫 で の み
1 2 3 4 5 6 7 8 9 1 0 11 1 2 1 3 1 4 1 5 16
"
図 - 6
( 1 ) 植物体及び媒介虫 の 全 DNA 試料か ら MLO に特異的 な プ ラ イ マ ー セ ッ ト を 用 い て PCR 法 に よ り 増 幅 し た DNA の ゲ ル泳動写真 ( 矢 印 ・ 750 bp) (H 健全試料 : D : そ れ ぞれの MLO に感染し た 試料)
1 λHind III
•
Eco RI マ ー カ ー2 タ マ ネ ギ ・ ツ ワ プ キ ・ イ ネ ( H ) の そ れ ぞ れ の PCR 反応試料 を 混合 し て 泳動
3 タ マ ネ ギ (D) 4 ツ ワ プ キ (D) 5 イ ネ (D)
6 Mycoþlasma orale 7 無議 ヒ メ フ タ テ ン ヨ コ バ イ
8 保議 ヒ メ フ タ テ ン ヨ コ バ イ
(2) 植物体及び媒介虫 の 全 DNA 試料か ら ユ ニ バ ー サ ノレ プ ラ イ 7 ー セ ッ ト 及 び MLO に特異的な プ ラ イ マ ー を 用 い て RPCR 法 に よ り 増 幅 し た DN A の ゲ ノレ泳動写真 (矢印上 1370 bp
: 下
750 bp) .9 λ H川d III • Eco RI マ ー カ ー
1 0 タ マ ネ ギ ・ ツ ワ プ キ ・ イ ネ ( H ) の そ れ ぞ れ の PCR 反応試料を混合 し て 泳動
11 タ マ ネ ギ (D) 1 2 ツ ワ プ キ (D) 1 3 イ ネ (D) 14 Mycoþ/asma 0叩le 15 無毒 ヒ メ フ タ テ ン ヨ コ バ イ 1 6 保毒 ヒ メ フ タ テ ン ヨ コ ノ マ イ
750 bp の DNA バ ン ド が増 幅 さ れ た 。 こ の DN A は , シ ー ク エ ン ス に よ り , I 群の MLO の 16S rRNA と 配列 が 一致 し た 。 以上の こ と か ら , 無毒虫 よ り 増幅 さ れた DNA は 恐 ら く ヒ メ フ タ テ ン ヨ コ パ イ に 寄 生 あ る い は共生 し て い る 微生物 由来の も の と 考 え ら れた 。 以上 の 結果 か ら , MLO 特異的 な プ ラ イ マ ー セ ッ ト を 用 い る こ と に よ り , 昆 虫 に 寄生 あ る い は 共生す る 微生物 を 保毒 す る M LO 無 毒媒介虫 と MLO 保毒媒介虫 と を 区別 す る こ と が で き る も の と 考 え ら れ る 。 し た が っ て , MLO 以外の微生物 と 相 向性の 高 い保存領域 を 有 す る プ ロ ー プ を 用 い た ハ イ プ リ
ダ イ ゼー シ ョ ン に よ る MLO 検 出 法 に 比 べ, 本 法 は 非 特 異反応が少な く 優れ て い る と い え よ う 。お わ り に
PCR 法 は , ( 1 ) 遺伝子 (DNA) を検出 す る 現在最 も 感 度 の 高 い 方法 で あ り , (2) 試料 と し て 使用 す る DNA の 純度 ( タ ンパ ク 質 . RNA な ど の 不純物の コ ン タ ミ ネ ー シ ョ ン の 問題) や DNA の サ イ ズ な ど は 余 り 問題 に な ら な い。 ま た , (3) 適切 な プ ラ イ マ ー を 使用 す れ ば, 操作は 簡単 で, 高 い再現性が得 ら れ, (4) 検出 ま で に 要 す る 時 聞が短い。 MLO は培養不能で あ り , 飾部局在性で あ る こ と か ら , 組織内 の濃度が き わ め て 低 < , 通常の 方 法 で は MLO の 16S rRNA 遺伝子 を 得 る の は 困難で あ り , 煩雑 であ る 。 し か し な が ら PCR は , 特異的 な プ ラ イ マ ー を 用 い て サ イ ク jレ条件 を 適当 に コ ン ト ロ ー ル す れば, 真核細 胞内の核及び、細胞内小器官の DNA を 除 か な く と も 細菌
O O iべE
PCR (Step-II)
MLQ genomlc DNA
\ミーJ
'
、唱』
--3
』、ー一ー一一『
図 - 7
RPCR 法の増幅反応、の手順 と 原理 を 説明 し た 図一一一
44一一一
16S rRNA遺伝子の PCR靖幅 に よ る 植物病原MLOの検出 と 分類 93
の 16S rRNA 遺伝子 を 得 る 便利 な 方法 と な る 。 本研究で
は, 酵素処理 に よ り 精製 し た MLO 分画だ け で な く , 植物 体 よ り 直接精製 し た 全 DNA か ら も , PCR 法 を 用 い る こ と に よ り , 容易 に MLO の 16S rRNA 遺伝子が増幅 さ れ る こ と が確認 さ れた 。 こ の遺伝子 は ま た , 媒介虫体内か ら も 増幅 さ れ, 他の共生 あ る い は 寄生性の細菌 と 区別す る こ と がで き た 。 ま た , MLO の 各群 に 特異的 な PCR プ ラ イ マ ー を 用 い る こ と に よ り , そ れ ぞ れ の 群 の 16S rRNA 遺伝子が特異 的 に 増幅 さ れ る こ と が確認 さ れた 。 さ ら に , 新 た に 二段階 PCR 法 を 考案 し , 1 本の チ ュ ー プ で複数の PCR 反応 を効率的 に し か も 経済的 に 行 う こ と がで き , MLO の所属群 を 特定 す る こ と がで き た 。 こ の 手 法 は , 今後他の分野 に も 応用 で き る も の と 考 え ら れ る 。 な お , 筆者 ら は現在, ア ジ ア 地域 に お け る MLO の 16S rRNA 遺伝子 の デ ー タ ベ ー ス 化 を す す め て お り , 将来,
こ れ ら の 成果か ら , MLO の遺伝子診断や, 分子分類への 道が開 け れ ば と 考 え て い る (NAMBA et
al.,submitted) 。 本研究 を行 う に 当 た り , 農水省農研セ ン タ ー の岩波節 夫, 加藤昭輔, 蚕見研の佐藤 守, 川 北 宏の 各氏 に は 様々 な ご援助 を い た だ い た 。 こ の 場 を借 り て 厚 く お 礼 申
し上 げ る 。
引 用 文 献
1) AHRENS, U. and SEEMÛLER, E. (1992) : Phytopathology 82 : 828�832.
2) CLARK, M. F., MORTON, A. and Buss, S. L. (1989) 目 Ann. Appl. Biol. 114 : 111�124.
3) DAVIS, R. E., LEE,
l.
.M., DoUGLAS, S. M. and DAI.LY, E.L. (1990) : Phytopathology 80 : 789�793.
4) DENG, S. and HIRUKI, C. ( 1991 ) : J. Microbiol. Methods 14 : 53�61.
5)
---
(1988) . " イ コ プ ラ ズ マ と そ の 実験法 (輿水 ら 編) . 近代出版 :287�306.6) 土居養二 ら ( 1967) : 日 植病報 33 : 259�266.
7) FELSENSTEIN, J. ( 1985) : Evolution 39 : 783�791.
8) GUTELL, R. R. et al. ( 1985) : Prog. Nucleic Acid Res Mol. Biol. 32 : 155�216.
9) G
凸 B
EL, U. B. and STANBRI郎E, E. J. (1984) : Science 226 : 1211 � 121310) 一一一一一 et al. (1987) : J. Gen. Microbiol. 133 : 1969
�1974.
11) HARASAWA, R., MIZUSAWA, H. and KOSHIMIZU, K.
(1986) : Microbiol. Immunol. 30 ・ 919-921 12) INNIS, M. A. and GELF
A
ND, D. H. ( 1990) : PCR Protocols : A Guide to Methods and Applications. (eds目 Innis, M. A. et aI.) . Academic Press, California,
pp. 3�12.
13) 加藤昭輔 ・ 岩波節夫 (1990) . 日 植病報 56 : 394 (講演要 旨)
14) ら (988) : 日 植病報 54 : 220�223 15) KIMURA, M. (1980) : ]. Mol. Evol. 16 : 11 1�120.
16) KIRKPATRICK, B.
c.
(991) : The prokaryotes 2nd ed.(eds. BALOWS, A. et aI.) . Springer-Ver\ag Press,
New York, pp. 4050�4067
17)
---
and FRASER, ]. D. (1988) : Phytopathology 79 : 1 138 (abstr.l18)
ー←一一一
et al. (1987) : Science 238 : 197�200.19)
一一一一一-
et al. (1990) : 8th 10M Congr. : 45�46 (abstr.) .20) KOLLAR, A. et al. ( 1990) : Phytopathology 80 : 233
�237.
21) KUNITA, S. et al. (1989) : Exp. Anim. 38 : 215�219 22) KUSKE,
c.
R. and KIRKPATRICK, B. C. (1990) ・ J. Bacteriol. 172 : 1628� 1633.
23)
一一一一一
(1992) : Int. ]. Sys. Bacteriol. 42 : 226�233.24) --- et al. (1991a) : J. Gen. Microbiol. 137 ・ 153
�159
25)
一一一一-
et al. (1991b) : Mol. Plant-Microbe Interact. 4 : 75�80.
26) LEE,
l.
-M. et al. (990) : Appl. Environ_ Microbiol.56 : 1471 � 1475.
27)
一一一一一一
and DAVIS, R. E. ( 1 986) : Annu. Rev.Phytopathol. 24 : 339�354.
28) LIM, P. ーO. and SEARS, B. B. (1989) : J. Bacteriol. 171 : 5901�5906.
29) LIN, C. P. and CIIEN , T. A. (1985) : Science 227 : 1233
�1235.
30) Lo, S. -C. et al. (989) ・ Am. J. Trop. Med. Hyg. 41 586�600.
31) MANILOFF, ]. (1983) : Annu目 Rev. Microbiol. 37 ・ 477
�499.
32) McCoy, R. E. et al. (1989) : The Mycoplasmas. Vol
ume V. (eds目 Whitcomb, R. F. et aI.) . Academic Press, New Y ork, pp. 545�640
33) 宮原和夫 ら ( 1982) : 日 植病報 48 : 551�554 34)
N A
KA
SHIMA
, K. et al. (1992) : Appl. Environ. Microbiol. 57 : 3570�3575
35) 奈須壮兆 ら (1967) 日 植病報 33 : 343�344 (講演要 旨) . 36) NEEFS, J. M. et al. (1990) : 1、�ucleic Acids Res. Seq.
Suppl. 18 : 2237�2317
37) NEIMARK, H 目 C. and LANGE, C.
s.
(1990) : Nucleic Acids Res. 18 ・ 5443�544838) RAZIN , S. (1985) : Microbiol. Rev. 49 : 419�455.
39) ROGERS, M. J. et al. (985) : Proc. N atl. Acad. Sci . U S. A. 82 : 1 160� 1164.
40) SAIKI, R. K. et al. (1988) : Science 239 : 487�491 41) SAITOU, N . and NEI, M. (1987) : Mol. Biol. Evol. 4 :
406�425.
42) SAWADA, M. et al. (1981) : Mol. Gen. Genet. 182 : 502
�504.
43) SEARS, B. B. et al. (1989) : Mol. Plant-Microbe Inter
act. 2 : 175� 180.
44) SoMMER, R. and T
A
UT
Z, D. ( 1989) : Nucleic Acids Res. 17 : 6749.45) TABOR, S. and RICIIARIlSON, C. C. ( 1989) : Proc. Natl.
Acad. Sci. U. S. A. 86 : 4076�4080.
46) WEISBURG, W. G. et al. (1989) : J. Bacteriol. 171 6455�6467.
47) WHITCOMB, R. F. (1980) : Ann. Rev. Microbiol. 34 677�709
48) WOESE,
c.
R. ( 1987) : Microbiol. Rev. 51 : 221�271.49) 一一一一一- et al. (980) : Proc. N atl. Acad. Sci. U. S.
A. 77 : 494�498.